ТРАНСПИРАЦИЯ
Вода — главный компонент живых растительных клеток, на ее долю приходится до 90% сырой массы. В зрелых растительных клетках большая часть воды содержится в крупной центральной вакуоли (Гэлстон, 1983), которая плотно прижимается к клеточной стенке, способствуя поддержанию формы клетки в мягких органах растения (листьях, стеблях, побегах). Форма декоративных травянистых растений зависит от гидростатических сил, при ослаблении последних вследствие потери влаги возникают утрата тургора и увядание. Если процесс увядания, например при сухом хранении срезки, не зашел далеко, при получении влаги запас воды в вакуоли возобновляется, тургор восстанавливается. Растения за счет специальных механизмов саморегуляции способны поддерживать свою оводненность на постоянном уровне, непрерывно поглощая воду и испаряя ее.
Транспирация представляет собой выделение растением водяного пара в атмосферу в основном посредством листьев и лепестков. Вода испаряется с поверхности влажных клеток, диффундирует по межклетникам и выходит наружу через открытые устьица либо кутикулу. Способность воды диффундировать, испаряться или поглощаться оценивают величиной водного потенциала, имеющего размерность давления. В отсутствие противодавления клеточной стенки поступление воды в клетку определяется водным потенциалом клетки, первоначально равным осмотическому потенциалу раствора, заполняющего вакуоль. С проникновением воды в вакуоль объем последней увеличивается, а раствор становится более разбавленным. Цитоплазма прижимается к стенке клетки, возникает противодавление клеточной стенки на клеточное содержимое. Когда противодавление клеточной стенки достигает достаточно большой величины, устанавливается динамическое равновесие, и приток воды прекращается.
Водный потенциал клетки определяется суммой потенциала давления и осмотического потенциала, который всегда отрицателен. Потенциал давления в живых клетках положителен. Клеточный потенциал равен нулю (при полном тургоре) или отрицателен при потере влаги и тургора. Если потенциал давления или тургорный потенциал упал до нуля, лист полностью завядает. Поэтому о завядании и потере влаги листьями и лепестками обычно судят по легко измеряемой величине водного потенциала соответствующих клеток (см. главу 8).
Интенсивность транспирации зависит от температуры и влажности воздуха, ширины раскрытия устьичных щелей, числа устьиц, облиственности срезки и разности водных потенциалов воздуха внутри и снаружи листа. Обычно вода из межклетников диффундирует в более сухой атмосферный воздух, и продолжительность сохранения срезки цветов в свежем состоянии определяется тем, как обеспечен транспорт к цветку воды и метаболитов, необходимых для поддержания жизнедеятельности.
В срезке цветов передвижение веществ по цветоносу отличается от аналогичного процесса на материнском растении, поскольку цветонос с цветком отделен от корней и частично листьев. Избыток воды, куда погружен цветонос, не может компенсировать активности корневой системы, обеспечивающей поступление веществ из почвенного раствора и веществ, синтезированных корневой системой, в том числе различных метаболитов и гормонов. Срезанный цветок в лучшем случае может использовать лишь то, что растворено в питающей его воде, засасываемой через срез цветоноса вследствие транспирации. Некоторые растворенные вещества, в частности сахара, адсорбируются плазматическим содержанием ситовидных трубок флоэмы, как это происходит и при движении синтезированных самим растением метаболитов. Вода перемещается по сосудам ксилемы цветочного побега, а тургор цветка и побега зависит от содержания в них воды.
Цветы различного анатомического строения, с неодинаковой сосудистой системой должны различаться и по срокам жизни в вазе. К числу наиболее устойчивых к завяданию в срезке декоративных растений относится тюльпан, наименее устойчивых — георгина. Промежуточное положение занимает душистый горошек. Гидравлическая характеристика сосудистой системы срезанных цветов определяется следующими факторами: отношением общей площади сосудов ксилемы к площади поперечного сечения стебля; средним количеством сосудов на один срез и на единицу площади срезки.
Существует прямая зависимость между способностью срезанного цветочного побега транспортировать воду и количеством сосудов в стебле, в то время как величина поперечного сечения сосудов (диаметр) играет прямо противоположную роль. У тюльпана средний диаметр сосудов меньше, но число их больше; у георгины — наоборот. Душистый горошек занимает промежуточное положение. Срезка тюльпана обладает наибольшей способностью к быстрому и полному восстановлению тур-гора, в то время как георгины даже после незначительного снижения тургора восстанавливают его с трудом, а при сильной потере тургора вообще не могут возвратиться в тургосцентное состояние, хотя сосуды георгины занимают примерно в 3 раза большую площадь, чем сосуды тюльпана. Более узкие и многочисленные сосуды ксилемы тюльпана обеспечивают лучшее водоснабжение цветка и стебля за счет капиллярных сил, величина которых тем больше, чем меньше диаметр трубок ксилемы.
Водный стресс в срезке обычно рассматривают как причину быстрого увядания вследствие закупорки проводящих сосудов стебля, которая может возникать в результате деятельности микроорганизмов либо под влиянием различных физиологических факторов. Баланс воды поэтому — один из главных факторов продолжительности жизни срезанных цветов. Со временем наблюдается снижение поглощения воды растением, сопровождающееся и уменьшением транспирации.
На рисунке 1 представлены кривые изменения транспирации и поглощения воды срезкой розы. Изменения в характере транспирации срезки роз сопровождаются изменением массы цветка. Причем это изменение неодинаково для цветов с длительным сроком жизни (роза Супер Стар) и более коротким (Голден Вэйв). В первые два дня наблюдается увеличение массы цветков и того и другого сорта, причем розы Супер Стар еще три дня сохраняют повышенную массу и лишь затем наблюдается ее снижение до уровня, все-таки превосходящего начальный. Поглощение воды превосходит транспирацию в течение первых пяти дней, затем эти показатели выравниваются и спустя примерно два дня транспирация начинает превосходить поглощение воды. Розы Голден Вэйв теряют массу начиная с третьего дня и уже на четвертый день их масса становится меньше первоначальной. Величины поглощения воды и транспирации монотонно снижаются в течение всего периода наблюдений. Измерения водопроницаемости (гидравлической проводимости) сосудистых каналов стебля указывают поначалу на резкое, а затем постепенное уменьшение этой величины для обоих сортов роз. Спустя 100 ч проводимость составляет около 0,2 первоначальной. Необходимо отметить, что проводимость сосудистой системы у свежесрезанных цветов не зависит от развития и возраста цветка.
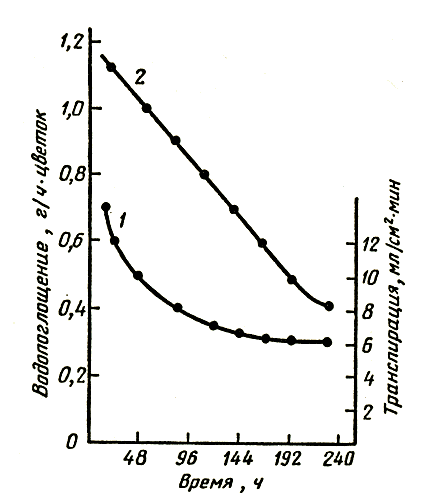
Рис. 1. Водопоглощение и транспирация срезки розы: 1 — транспирация; 2 — водопоглощение
Водный потенциал лепестков резко уменьшается спустя примерно шесть дней, чем и определяется продолжительность жизни цветов в вазе. Такое снижение водного потенциала происходит не вследствие естественного старения, поскольку непосредственно на вегетирующем растении уменьшения водного потенциала в течение того же времени не наблюдается. Раннее старение цветочной срезки вызывается развитием водного стресса, ведушего к снижению соответствующего потенциала, причем этот процесс происходит более быстро у сортов с коротким периодом жизни.
Закупорка сосудов — основная причина увядания и старения срезанных цветов, что подтверждается снижением гидравлической проводимости стебля, одинаковым для обоих сортов. Это снижение проводимости нельзя связать с естественным старением, поскольку у вегетирующего растения она не изменяется, уменьшение проводимости происходит в результате отделения цветка от материнского растения.
Повышение со временем ферментной активности у срезанных цветов прямо указывает на то, что закупорка сосудов связана с этим явлением, а не только с активностью микрофлоры. Под влиянием ферментов происходит гидролиз стенок клетчатки, выстилающих сосуды, их гидравлическое сопротивление увеличивается. Поэтому один из возможных способов продления срока жизни срезанных цветов — инактивация ферментов.
Характерно, что как повышение ферментной активности, так и снижение проводимости сосудистой системы примерно одинаково и для цветов с коротким, и с длинным периодами жизни в вазе. Различие в этих сроках можно объяснить исходя из водного баланса в срезке, определяемого количеством поступившей воды и воды, потерянной цветком в результате транспирации. У цветков с коротким периодом траспирация выше, водный потенциал ниже, что и приводит к более раннему увяданию. Эта разница в транспирации — следствие различной способности устьиц реагировать на стрессовую ситуацию. На материнском растении раскрытие устьиц одинаково для обоих сортов, но после срезки раскрытие устьиц у короткоживущего сорта Голден Вэйв больше. Если удалить у срезки все листья, то разница в потерях влаги и продолжительности жизни нивелируется. К этому же приводит и обработка специальными препаратами, вызывающими закрытие устьиц.
Тем не менее объяснить разницу в сроках жизни лишь на основе механизма, регулирующего транспирацию путем открытия устьиц, невозможно. Большую роль в этом играет и активность различных метаболитов. Увядание поставленных в воду цветов происходит значительно быстрее, чем тех же цветов на материнском растении, хотя по обеспеченности водой они могут быть сопоставимы. Это увядание связано с уменьшением Сахаров. в тканях цветов, поскольку добавление в воду сахарозы или глюкозы увеличивает срок жизни цветов примерно вдвое. Такие результаты были получены в опытах с луковичными растениями — тюльпаном, нарциссом и гиацинтом и с другими декоративными растениями — сиренью, душистым горошком.
Содержание воды в цветках душистого горошка в вазе на второй и третий день было выше, чем в цветках на растении. Затем содержание воды в них резко упало, и цветы завяли. Аналогичная картина отмечена и у тюльпана, причем одновременно происходило снижение интенсивности дыхания.
Таким образом, положительное влияние на продление срока жизни в воде таких цветов, как тюльпан, нарцисс, гиацинт, ландыш, душистый горошек, оказывают вещества, повышающие или стабилизирующие интенсивность дыхания.
Иначе обстоит дело с транспирацией и передвижением воды у декоративных растений, цветы или соцветия которых быстро теряют тургор после срезки (даже если их незамедлительно помещают в воду), хотя на материнском растении они держатся долго. Характерный представитель этой группы растений — георгина. Увядание соцветий у нее связано с постепенной закупоркой сосудов, увеличением их гидравлического сопротивления, что приводит к нарушению транспирации и обезвоживанию тканей цветка. Это, в свою очередь, ведет к нарушению ферментативного равновесия и гидролизу белков плазмы вплоть до гибели растения. Интенсивность дыхания цветков георгины в вазе снижается резко и лишь при увядании отмечается климактерический подъем дыхания. Добавление в воду сахарозы или глюкозы не влияет на продолжительность жизни в вазе цветов георгины, но зато большое влияние оказывают вещества, способствующие передвижению по сосудам и препятствующие их закупорке (сернокислый алюминий, окисное сернокислое железо и некоторые другие).
Следовательно, различные виды декоративных растений нуждаются в специфических веществах для продления срока жизни в вазе. Растения с высокой интенсивностью дыхания для поддержания ее на должном уровне и цветы, интенсивность дыхания которых в вазе быстро падает, испытывают потребность в сахарах. На растения, сосудистая система которых имеет тенденцию к закупорке, положительное влияние оказывает введение в воду веществ, способствующих лучшему передвижению питательного раствора в тканях цветка.
Применение одних и тех же веществ, рекомендуемых для продления срока жизни цветов в вазе, в одних случаях дает положительный эффект, в других — никакого, в подчас получают даже отрицательный результат. Более того, в одних местностях срезка в обычной воде стоит дольше, чем в других, что указывает на различное качество воды в отдельных регионах. Смягчение жесткой воды с помощью соды, как правило, приводит к ухудшению условий содержания срезки гвоздики — длительность жизни цветов в вазе снижается. Качество воды оказывает влияние и на раскрытие бутонов гвоздики, срезанной в период полуроспуска, повышенное содержание минеральных солей в воде задерживает этот процесс. Растворенные в воде фторсодержащие соединения могут вызвать некроз лепестков и листьев срезки гладиолуса. Некоторые цветы имеют особое анатомическое строение стебля, представляющего собой полую трубку, в стенках которой располагаются проводящие сосуды. Такое строение имеют цветы герберы (рис. 2), у которой потеря декоративных качеств выражается в «поникающей головке», то есть в потере тургора стеблем при относительно свежем цветке. Измерение водного потенциала лепестков у поникших цветков показало, что он достаточно высок и близок к потенциалу незавядшей срезки герберы, в то время как стебель испытывает явный водный стресс (его влагосодержание сильно уменьшилось), по-видимому, лепестки герберы обладают способностью отбирать воду от стебля.
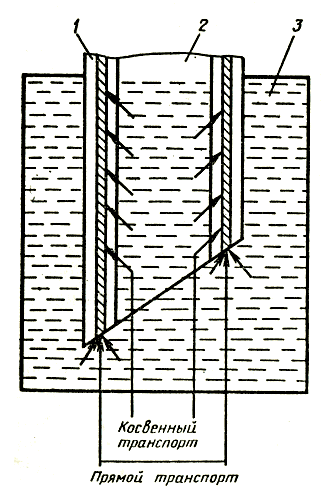
Рис. 2. Строение стебля герберы: 1 — полость в стенке цветоноса; 2 — полость внутри цветоноса; 3 — вода вазы
Водный баланс герберы складывается из водопоглощения и транспирации. После срезки транспирация остается примерно на том же уровне, а водопоглощение монотонно падает. Последнее определяется градиентом водного потенциала и гидравлическим сопротивлением на дистанции ваза — лепестки. Уменьшение водопоглощения может быть вызвано как снижением градиента потенциала, так и увеличением гидравлического сопротивления. Поскольку градиент потенциала изменяется незначительно, уменьшение водопоглощения связано с увеличением гидравлического сопротивления, что отмечено и для некоторых других цветов, например розы.
Чтобы предотвратить потери тургора стеблем, можно использовать различные химические вещества, имеющие бактерицидное действие. Эти вещества предотвращают и микробиологическую закупорку сосудов ксилемы, блокируя прямой водный транспорт. Однако, если вода попадает в полость стебля, возможен косвенный транспорт ее через ткани стенки стебля. Поэтому для цветов, имеющих такое же строение стебля, как у герберы, на некотором расстоянии над поверхностью воды в вазе нужно сделать небольшое отверстие в стебле для выпуска воздуха. В этом случае срезанные цветы стоят на 5—10 дней дольше.
Водопоглощение в срезанных цветах зависит также от неразрывности водного пути между вазой и растением, который может блокироваться воздушными тромбами в проводящих сосудах ксилемы. Эти воздушные тромбы обычно возникают в виде небольших пузырьков на стенках или в углублениях сосудов, и для хорошей сохранности срезки необходимо как можно быстрее восстановить целостность транспортирующих потоков ксилемы.
При срезывании цветоносов вода выдавливается из кончика срезанного стебля и образуется воздушный тромб, величина которого зависит от водного потенциала цветка до срезывания и интенсивности транспирации после этого. Наличие таких тромбов препятствует нормальной транспирации, водный поток устремляется в обход тромба через ткани, окружающие сосуды ксилемы, что увеличивает гидравлическое сопротивление в десятки раз. Повторная обрезка стебля под водой устраняет воздушную блокировку сосудистых пучков ксилемы. Аналогичный результат дает и добавление в воду, где находится срезка цветов, некоторых подкисляющих, смачивающих или поверхностно-активных веществ, способствующих рассасыванию воздушных тромбов. Если вследствие закупорки сосудов вода не может подниматься по стеблю, для облегчения тока воды по сосудистой системе можно использовать устройство, подающее воду в срезку под давлением. Подробно этот метод рассмотрен в главе 3.
Водный поток через ксилему, гидравлическая проводимость ее сосудов и транспирация взаимосвязаны. Объемный расход (м3/с) воды через сосуды ксилемы можно определить по формуле
Q - PPS, (4)
где р — гидравлическое давление в сосудах ксилемы, Па; Р — гидравлическая проводимость; S — площадь поперечного сечения сосудов, м2.
Отсюда следует, что транспирационный поток при прочих равных условиях зависит от проводимости сосудов ксилемы. Поскольку водный ток обеспечивает связь между стеблем и цветком, весьма важно не допустить блокировку сосудов ксилемы, повышения ее гидравлического сопротивления, чтобы не уменьшить насыщенность водой (тургор) лепестков и предотвратить быстрое увядание. Следует также иметь в виду, что транспирационный ток воды обеспечивает протекание процессов обмена, связь срезки с питательным раствором вазы. Жизнь срезанных цветов в вазе будет тем продолжительнее, чем лучше работает транспирационный ток ксилемы, дающий срезке необходимое количество влаги.
Количественно транспирацию оценивают величиной потока, отнесенного, как правило, не к площади сосудов ксилемы, поскольку эта величина трудно определима, а к площади поверхности листа или листьев. Такое определение транспирации дает возможность проводить сопоставимый анализ движения воды в сходных растениях. При этом, однако, возможны систематические ошибки, поскольку не всегда поверхности испарения сопоставимы друг с другом: устьица могут располагаться как с одной, так и с обеих сторон листа и т. д. Кроме того, точное определение площади листьев также представляет трудности.
Для того чтобы возместить потери воды срезкой при испарении, расход воды на испарение должен соответствовать приходу, зависящему от транспирационного потока через сосуды ксилемы. При отсутствии такого баланса например в результате закупорки части сосудов, может возникнуть водный дефицит, вызывающий физиологические расстройства срезки и ее гибель.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://flowerlib.ru/ 'Библиотека по цветоводству'