ФИТОГОРМОНЫ И ЭТИЛЕН
Завядание цветка как на материнском растении, так и в срезке — одна из форм старения растительных тканей. Между старением растительных тканей и содержанием в них ауксинов и ингибиторов роста имеется определенная связь. Опадение бутонов, иногда наблюдаемое в срезке, вызывается нарушением баланса между ауксинами и ингибиторами, и в стареющих тканях происходит замедление передвижения ауксинов и цитокининов. Продление жизни срезанных цветов в вазе до тех же сроков, что и в естественных условиях на материнском растении, возможно при добавлении к воде со срезкой сахарозы или глюкозы, стабилизирующих дыхание и метаболические процессы в растении. Большая роль в этом процессе принадлежит регуляторам роста, в частности эндогенным ауксинам и хлорогеновой кислоте, что было подтверждено опытами со срезкой тюльпанов, находящихся в дистиллированной воде и в 6%-ном растворе сахарозы.
Вначале, при появлении окраски в бутонах (первый день), количество ауксинов в околоцветнике и пестике одинаково как в срезке, так и в цветках на растении. Затем вместо характерного для материнского растения резкого возрастания количества ауксинов наблюдается противоположное явление — снижение их содержания. При этом уменьшение количества ауксинов остается необратимым. Добавление к воде сахарозы вызывает (по сравнению со срезкой в чистой воде) повышение содержания ростактивирующих веществ, снижение количества ингибиторов роста. Это задерживает завядание — околоцветник в варианте с чистой водой осыпается к восьмому дню, в то время как у цветков в растворе сахарозы остается свежим, сохраняя свои декоративные качества. Действие сахарозы связано с увеличением интенсивности дыхания цветка, способствующим повышению уровня его жизнедеятельности, направленной на усиление синтеза ауксинов при снижении синтеза ингибиторов. Такое изменение между регуляторами роста интенсифицирует перемещение в срезанный цветок метаболитов, еще сохранившихся в тканях цветочного побега.
Аналогичная картина наблюдается и в отношении кинетики содержания ингибитора некоторых ферментов — хлорогеновой кислоты. Добавление сахарозы способствует усилению синтеза хлорогеновой кислоты, что изменяет соотношение между регуляторами роста, приближая состояние срезанного цветка к физиологическому его состоянию на растении.
К фитогормонам, вырабатываемым как здоровыми клетками растений, так и в особенности клетками, подвергшимися физиологическому разрушению, относится этилен. Известно, что одно гнилое яблоко в контейнере вызывает порчу остальных. Это связано с тем, что в нем вырабатывается летучий газ — этилен, вызывающий разрушительные изменения в соседних здоровых плодах. Плоды портятся и, в свою очередь, начинают выделять этилен, в результате чего возникает цепная реакция порчи.
Среди эндогенных регуляторов метаболических процессов, таких как созревание плодов или опадение листьев, этилен вследствие его летучести занимает особое место. В отличие от других гормонов транспорт этилена не требует каких-то специализированных механизмов и осуществляется практически беспрепятственно. Чтобы устранить избыточное количество фитогормонов, нет нужды и в специальных системах инактивации — газ выводится в окружающее пространство. Некоторые проблемы возникают лишь тогда, когда это пространство ограниченно, например объем хранилища. Такое своеобразие свойств этилена обусловило довольно позднее признание его в качестве регулятора метаболических процессов, а не как сопутствующий этим процессам побочный продукт.
Этилен — ненасыщенный углеводород (С2Н4), представляет собой бесцветный газ с запахом, напоминающим запах диэтилового эфира. Этилен легко вступает в реакцию с галогенами, водой, серной кислотой, легко окисляется кислородом, озоном, перманганатом калия. Способность этилена инактивироваться под воздействием окислителей имеет большое значение в технологии хранения плодов, овощей, цветов. Добавление марганцовокислого калия в герметизированный контейнер предотвращает накопление этилена и порчу срезки цветов или посевного материала (семена, луковицы, клубнелуковицы и т.п.). Этой же цели служит и обработка озоном с помощью, например, барьерного разряда (см. главу 3).
Для этилена характерна высокая активность при очень малых концентрациях (единицы или десятки мкл/л). Выявление этилена в таких концентрациях возможно лишь чувствительными методами хроматографического анализа.
Определенным антагонистом этилена служит углекислый газ, причем соотношение между этими компонентами таково. Чтобы нейтрализовать активность одной молекулы этилена, необходимо 105 молекул углекислого газа. Хранение цветов в модифицированной среде, обогащенной углекислым газом, дает возможность нейтрализовать действие продуцируемого растениями этилена, если концентрация углекислого газа достаточно высока. Этилен способен быстро перемещаться по клеточным тканям и свободному межклеточному пространству — при обработке этиленом одного листа его активность быстро проявляется во всех органах растения. Перемещение этилена происходит практически беспрепятственно, с чем и связана его способность выполнять функции посредника в процессах метаболизма с участием фитогормонов.
Этилен вызывает старение клеток, тканей и органов, тормозит деление клеток. Весьма велика роль этилена, индуцированного повреждением растения. Такой стрессовый этилен образуется при экстремальных температурных воздействиях, ухудшении водоснабжения, при заболеваниях, вызванных бактериями и грибами, при механических травмах, поскольку стрессовый этилен способствует отторжению поврежденных тканей и органов.
Инактивация как эндогенного, так и стрессового этилена имеет большое значение в технологии хранения. Как срезка, так и материал для размножения растений (семена, луковицы, клубни) выделяют этилен, ускоряющий созревание и сокращающий сроки хранения. Повышение концентрации углекислого газа и снижение содержания кислорода способны резко замедлить синтез этилена. Этим путем можно обспечить более продолжительное хранение срезки и семенного фонда, особенно в условиях пониженных температур.
Физиологическое действие этилена определяется тремя главными факторами: концентрацией, длительностью воздействия и температурой. Иногда целесообразно рассматривать воздействие дозы этилена, которая представляет собой произведение концентрации на продолжительность его действия во времени. Такой подход наиболее удобен при изучении действия этилена на срезку цветов, например гвоздики (Barden, 1972). На рисунке 3 представлена зависимость относительного срока хранения срезки гвоздики Уайт Сим от дозы этилена. Предварительно было установлено, что этилен обладает кумулятивным действием — один и тот же эффект увядания наблюдается при концентрации 0,4 мкл/л в течение 10 ч и 0,04 мкл/л в течение 100 ч.
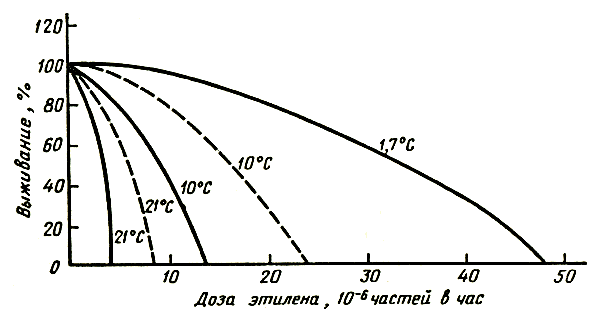
Рис. 3. Влияние этилена на сроки хранения гвоздики Уайт Сим: сплошная линия — срезка; пунктирная — черенки
Анализ кривых, приведенных на рисунке, свидетельствует о том, что гвоздика в бутонах лучше сопротивляется действию этилена, чем распустившиеся цветы. Весьма существенно и действие температуры. Раскрывшиеся цветки гвоздики в 1000 раз более чувствительны к этилену при повышении температуры с 1,7 до 21°С. Гвоздика в полуроспуске в 10 раз более чувствительна к действию этилена при повышении температуры с 10 до 21°С.
Какова же предельно допустимая концентрация этилена с точки зрения технологии хранения? Очевидно, что ответ на этот вопрос связан с предельно допустимой нормой отхода срезки при хранении. Считается, что эта величина составляет примерно 20%. Тогда можно получить зависимость между дозой действия этилена и температурой, соответствующей данной величине потерь (20%). При температуре 4°С предельно допустимая доза для раскрытых цветков будет примерно 10 мкл/л в час. При более высоких температурах эти предельно допустимые дозы быстро уменьшаются.
Для расчетов холодильных камер необходимо располагать сведениями о количестве этилена, продуцируемого цветами. Приводим данные исследователя Фан Кхон Тона о количестве этилена (мкл/кг в сутки), выделяемого срезкой цветов при 20°С:

|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://flowerlib.ru/ 'Библиотека по цветоводству'