ОБЩИЕ ОСНОВЫ ВЫГОНКИ ДЕКОРАТИВНЫХ ЛУКОВИЧНЫХ РАСТЕНИЙ
В соответствии с исторически сложившимся в процессе эволюции генотипом все виды существующих в природе растений цветут в определенные запрограммированные сроки, в той или иной степени корректируемые внешними условиями среды обитания. Точные даты цветения, присущие различным видам растений, достаточно точно определены ботаниками в процессе фенологических наблюдений, проводимых в различных природно-климатических зонах земного шара на протяжении ряда лет. Естественно, что для одних и тех же видов растений с широким ареалом расселения календарные сроки начала цветения могут различаться в очень больших пределах, достигающих иногда нескольких месяцев.
Особенно наглядно эти различия выявляются при сопоставлении сроков цветения одних и тех же видов, произрастающих в южных и северных районах одной страны и тем более в Северном и Южном полушариях.
На основе этой естественной способности растений цвести в различные сроки в соответствии с комплексом внешних условий в течение длительного времени строилась вся система снабжения живыми цветами населения во всех странах мира.
Поступление срезанных или горшечных цветов в продажу определялось прежде всего умелым подбором ассортимента цветочно-декоративных культур, обеспечивающим их живой конвейер в течение вегетационного периода.
Новые возможности в этом отношении появились в период небывалого расширения границ международного рынка и международных торговых связей, тесно соединивших между собой не только отдельные страны, но и континенты.
В этих условиях в самых широких масштабах стал практиковаться активный коммерческий обмен цветочной продукцией между различными районами земного шара, и доставка даже самых экзотических видов растений в необычное время туда, где о них раньше не могли и мечтать, не вызывала особого удивления.
Однако наиболее кардинальное решение проблемы снабжения населения цветами на протяжении круглого года было достигнуто после того, как в большинстве передовых европейских стран господствующее положение в цветоводстве заняли крупные оранжерейные хозяйства, освоившие индустриальные способы производства.
Массовое строительство современных оранжерей и крупных оранжерейных комплексов, вооруженных автоматикой, позволяющей регулировать, по существу, все основные показатели микроклимата внутри культивационных помещений, устранило, казалось бы, непреодолимые прежде препятствия на пути управляемой культуры большого числа декоративных растений. Появились реальные предпосылки по заранее составленной программе в нужных пределах изменять в оранжереях температурный режим, условия увлажнения, минеральное питание, а в последние годы и освещенность растений и тем самым конкретно управлять сроками их цветения.
Следует особо подчеркнуть, что научно-технический прогресс в области промышленного цветоводства был дополнен существенными достижениями в разработке более совершенных приемов размножения и выращивания растений, а в ряде случаев и целых технологий, основанных на глубоком знании закономерностей морфогенеза многих декоративных растений и выявлении физиолого-биохимических процессов, лежащих в их основе.
Независимо от местных природно-климатических условий, используя преимущества новых современных оранжерей и научно обоснованные рекомендации, производственные хозяйства получили возможность производить цветочную продукцию в заранее определяемые сроки.
Небывало широкое распространение в связи с этим в практике цветоводческих хозяйств и оранжерейных комплексов получил хорошо известный среди специалистов-цветоводов способ так называемой выгонки декоративных растений. Этот термин, давно получивший права гражданства в декоративном садоводстве, обозначает совокупность приемов воздействия на растение, обеспечивающих его цветение в необычные сроки. Так, на протяжении многих десятков лет создавались и совершенствовались широко известные системы выгонки (или выгоночная культура) роз и сирени, ремонтантной гвоздики и хризантем, герберы, ландышей и многих других цветочно-декоративных растений, создающих практически почти непрерывный конвейер живых цветов в течение года.
Наряду с конкретными принципиальными различиями, определяющимися биологическими особенностями растений, все указанные выгоночные системы, которые, по нашему мнению, на данном этапе с полным основанием должны рассматриваться как «управляемые культуры», имеют общие основы, общие аналогичные элементы, их составляющие.
Вне зависимости от культуры каждая система обязательно включает следующие неразрывно связанные между собой элементы технологии.
а. Отбор исходного сортового и посадочного материала, в значительной мере определяющий успех дальнейшей работы.
б. Предварительная подготовка, обработка посадочного материала перед посадкой (прогревание, охлаждение и т. п.).
в. Экспериментально проверенные сроки и способы посадки на выгонку, увязанные с графиком поступления цветущих растений.
г. Активные воздействия па рост и развитие растений различного рода факторами внешней среды (изменение температурного, водного, пищевого режимов, фотопериодическая индукция, обрезка, прищипка и др.) на соответствующих этапах онтогенеза с целью регулирования сроков цветения с учетом времени получения цветочной продукции.
Естественно, что технологией также предусматриваются обычные агротехнические меры по уходу за выгоночными растениями, способы срезки цветков и условия их хранения.
В связи с рассматриваемыми нами в данной работе основами выгонки цветочно-декоративных луковичных растений вполне уместно обратить внимание читателей на то, что в данном случае целесообразность замены термина «выгонка» на более точное понятие «управляемая культура» наиболее обоснована. Правомерность подобного утверждения определяется не только глубиной и обстоятельностью научной разработки технологии выгонки важнейших цветочных луковичных растений (тюльпаны, парциссы, гиацинты), но и тем, что все ее органически связанные между собой элементы прошли надежную экспериментальную и производственную проверку, подтвердившую высокую эффективность в самых различных условиях.
В современном виде управляемая культура цветочных луковичных растений возникла при участии ученых многих стран мира, однако основной вклад в разработку общебиологических основ и ряда важных приемов, ее составляющих, внесли многие поколения голландских цветоводов, а в последние годы научные сотрудники Лаборатории по луковичным растениям в г. Лиссе и института в г. Вгенингене.
Как известно, цветочные луковичные растения относятся к разным семействам, а их распространение приурочено к различным местообитаниям. Общность их заключается в том, что зачатки мопокарпического побега формируются внутри луковицы за счет веществ, накопленных в запасающих чешуях, и в течение значительного времени побег находится в условиях, в известной степени независимых от непосредственного воздействия внешней среды.
В этой связи изучение и познание общих особенностей морфогенеза, отдельных его этапов и морфологических структур, а также развития и коррелятивных связей подземных и надземных органов у луковичных растений на протяжении длительного времени привлекало внимание голландских ученых, ибо оно открывало возможность для сознательного изменения и управления процессами, происходящими в растении на соответствующих этапах онтогенеза. Начало исследованиям данной группы растений в этом направлении было положено Ирмишем [Irmisch, 1863]. Однако особенно интенсивные исследования луковичных культур развернулись в Голландии в 1920—1940-х годах.
Даже беглый просмотр опубликованных в этот период работ свидетельствует о том, что они проводились широким фронтом, охватывая наряду с морфогенезом обширный комплекс факторов, определяющих цветение растений. Особое место в исследованиях, относящихся к этому времени, занимают работы Блау (Blaauw), который вместе со своими сотрудниками в течение почти 20 лет провел обстоятельное изучение 23 родов луковичных, клубнелуковичных и некоторых клубневых цветочных растений.
Блау и его сотрудники сумели установить наиболее существенные закономерности в морфологическом формировании луковичного растения на различных этапах онтогенеза и определить циклы термопериодического оптимума, оказывающего влияние на органообразование внутри луковицы.
Особое внимание было обращено на образование зачатков цветка и цветоноса, изучены и зафиксированы все стадии дифференциации всех элементов цветка.
Впервые в 1926 г. Блау представил общую схему морфогенетических изменений в луковице, описав стадии заложения и формирования цветка у луковичных растений (тюльпаны), значительно облегчившую дальнейшую разработку научных основ выгонки. Отдельные стадии были обозначены римскими цифрами в зависимости от состояния зачатка цветка.
Позднее, в 1942 г. Бейджер [Beijer, 1942] ввел в эту схему морфогенеза луковичных растений буквенные обозначения отдельных стадий, используя первые буквы ботанической номенклатуры элементов цветка (Р — perianthium, A — androecium, G — gynoecium).
Ниже приводится схема морфогенеза, характеризующая стадии заложения и формирования цветка, предложенная Блау и Бейджером, получившая всеобщее признание.

Схема морфогенеза, характеризующая стадии заложения и формирования цветка, предложенная Блау и Бейджером, получившая всеобщее признание
Следует уточнить, что не все эти стадии обязательны для отдельных родов луковичных растений, но знание условных обозначений стадий развития почки возобновления весьма важно при освоении технологии выращивания и выгонки растений. В качестве примера укажем, что для луковичных растений семейства Амариллисовых Г. Е. Капинос установила 6 основных фаз развития, подразделенных на 10 этапов:
I фаза — Внутрилуковичное развитие:
1) недифференцированный конус нарастания,
2) дифференциация зачатков листьев,
3) заложение оси цветка, прицветных листьев и второй почки возобновления,
4) формирование органов цветка,
5) микро- и мегаспорогенез.
II фаза — Внелуковичный подземный рост почки возобновления:
6) усиленный подземный рост листьев.
III фаза — Вегетация:
7) появление листьев на поверхности земли, их рост и ассимиляция.
IV фаза — Бутонизация:
8) появление бутона над землей.
V фаза — Цветение:
9) раскрывание бутонов, массовое цветение.
VI фаза — Плодоношение:
10) формирование и созревание плода.
Возможно, с технологических позиций эта схема излишне подробна, но, несомненно, она дает четкую картину этапов морфогенеза растений данного семейства и может быть использована при решении многих вопросов, связанных с регулированием сроков цветения отдельных видов.
Существенный вклад в систематизацию накопленных к этому времени данных по морфогенезу луковичных растений внес Хартсема [Hartsema, 1961], уточнив их дифференциацию в зависимости от времени заложения и последующего развития цветка в луковицах в пределах 5 групп:
I группа — Цветки закладываются весной или в начале лета в год, предшествующий цветению, до уборки луковиц (нарциссы, подснежники, леукоюм).
II группа — Цветки закладываются после завершения периода роста, т. е. осенью после уборки (тюльпаны, гиацинты).
III группа — Цветки закладываются осенью после посадки или ранней весной (лилии).
IV группа — Цветки закладываются более чем за год до цветения (нерине).
V группа — Цветки закладываются в разное время вместе с образованием листьев в течение всего периода роста (гиппеаструм).
Подобное подразделение луковичных растений по столь важному для определения сроков воздействия на общий ход развития и формирование зачатков цветочного побега признаку имел важное значение в дальнейшем при разработке технологии выгонки отдельных культур.
Не менее результативными в этом отношении оказались исследования школы Блау, посвященные комплексному изучению морфологического состояния точки роста растения на различных этапах онтогенеза и влияния температуры на ее изменение.
Установление циклов термопериодического оптимума у тюльпанов и гиацинтов позволило сделать важнейшее открытие, убедительно подтвердившее, что для луковичных растений температура является самым ответственным фактором, оказывающим решающее влияние на заложение и последующее развитие цветка.
В последующем исследованиями Хартсемы было показано, что образование цветка у луковичных растений (тюльпаны, гиацинты) не связано ни с интенсивностью света, ни с продолжительностью дня, так как цветок закладывается в период «покоя». У гиппеаструма образование цветка проходит в течение всего года.
Многочисленными прямыми экспериментами было установлено, что для каждого рода луковичных растений существует свой режим температурной обработки, тесно связанный с определенными этапами морфогенеза и, конечно, со сроками выгонки в оранжерее.
Однако практически процесс температурной обработки луковиц, подготавливаемых для выгонки, как правило, подразделяется на два этапа, различающихся по условиям и продолжительности.
Первый этап связан с завершением формирования всех органов цветка. В это время используют повышенные температуры, ускоряющие этот процесс. Переход от образования листьев к стадии формирования цветка при искусственном создании оптимальных условий происходит очень быстро. В работах Камербека и Хугетерпа [Нооgeterp, 1969] было показано, что период от наступления стадии G и до момента цветения очень стабильный, т. е. сезонные условия мало отражаются на темпах развития цветочного побега в луковице. Однако наступление стадии G варьирует по сезонам и в какой-то мере зависит от времени уборки. Быстрое образование и развитие зачатков цветков в луковице положительно сказывается на раннем цветении в выгонке.
По данным указанных авторов, обработка высокими температурами стимулирует развитие зачатков листьев и дифференциацию точки роста, заложение элементов цветка идет быстрее, чем без обработки. В дальнейшем было установлено, что на первом этапе для оптимального завершения процесса дифференциации зачатка цветка для тюльпанов необходима температура 17—20°, нарциссов — 17°, гиацинтов — 25,5°, мускари, галантуса — 15—17°. Более высокая и низкая температуры задерживают этот процесс. Лишь при подготовке луковиц для ранней выгонки (в декабре, январе) прибегают обычно к их обработке с помощью повышенных температур сразу после уборки. В частности, для тюльпанов температуру повышают до 34° в течение 1 нед, а для нарциссов на 4 дня, луковицы гиацинтов в течение 2 нед хранят при 30°.
В известной мере «шоковая» температура несколько задерживает развитие листовых зачатков и стимулирует дифференциацию цветка.
После завершения первого этапа подготовки, когда зачаток цветка в луковице достигнет необходимой стадии (стадия G), наступает второй этап холодного хранения луковиц (при 9°), способствующий снятию покоя у растений. В естественных условиях луковичные растения находятся в состоянии покоя при низких температурах в течение 5—6 зимних месяцев после посадки.
В условиях хранилища при охлаждении луковиц с полностью сформированным цветочным побегом в столь длительном периоде охлаждения нет необходимости.
Установлено, что период охлаждения у тюльпанов при температуре 9° в зависимости от сорта составляет 12—24 нед, у нарциссов — 13—18 нед, у гиацинтов 10—12 нед. Слишком продолжительный период охлаждения стимулирует ростовые процессы и способствует снижению прочности стебля, искривлению и полеганию растений. При недостаточно продолжительном периоде охлаждения в выгонке образуются низкорослые растения, а некоторые виды и сорта не цветут вообще, образуя «слепые» бутоны. Любое отклонение от установленного режима охлаждения отрицательно сказывается на качестве цветущих растений, а при грубом нарушении его ведет к повреждению и гибели зачатков цветка в луковице. В связи с этим очень важно и при отборе посадочного материала до начала и в процессе выгонки контролировать заложение и формирование цветка в луковице. Для этого начиная с момента уборки следят за формированием цветка, систематически отбирая пробы. Ранние стадии образования зачатков цветочного стебля можно установить под бинокулярной лупой или микроскопом (при увеличении 40). Чтобы точно определить стадии дифференциации цветка, просматривают по несколько луковиц из всех партий посадочного материала, поступившего на выгонку, особенно если они были получены из различных хозяйств и разных природно-климатических районов.
С целью получения цветущих растений в течение всего зимне-весеннего периода выгонку луковичных культур (тюльпаны, нарциссы, гиацинты), как правило, проводят по партиям, различающимся по срокам выгонки:
I — в конце декабря — начале января,
II — в январе — начале февраля,
III — в конце февраля — начале марта,
IV — в конце марта — начале апреля,
V — в конце апреля — начале мая.
При планировании выгонки луковичных культур в определенные сроки в каждом конкретном случае следует учитывать следующие факторы: биологические особенности сортов; исходную массу и размер луковиц, температуру в период хранения после уборки луковиц из открытого грунта; время посадки на выгонку; температурный режим и влажность субстрата и воздуха в период укоренения и охлаждения; температурный режим в период выгонки и освещение растений.
Со всем этим комплексом факторов, определяющим успешность подготовки и выращивания растений, мы подробно ознакомимся в следующих главах, посвященных частным технологиям выгонки отдельных луковичных культур.
В заключение этой главы, освещающей общие биологические основы управляемой культуры цветочных луковичных растений, вполне целесообразно коротко остановиться на физиолого-биохимических закономерностях, определяющих процесс цветения растений при выгонке.
Это тем более необходимо, так как выяснение внутренних механизмов ростовых и формообразовательных процессов, несомненно, может иметь важное практическое значение для разработки методов управления жизнедеятельностью растительных организмов.
Фундаментальные исследования, проведенные физиологами за последнее десятилетие, сыграли важную роль в решении многих вопросов, связанных с выращиванием, размножением и выгонкой луковичных растений. К такого рода работам следует отнести исследования физиологических закономерностей морфогенеза, очень существенные для возможного в дальнейшем искусственного изменения и управления ходом этих процессов. По современным представлениям состояние покоя и рост растений регулируются фитогормонами и эндогенными ингибиторами [Чайлахящ 1988].
Особое значение в этом направлении приобретают исследования гормональной регуляции роста и морфогенеза у луковичных растений, проведенные сравнительно недавно в Голландии Перейра [Pereira, 1984], в США Оунгом, Хертогом [Aung, Hertogh, 1968a, b, 1979], в Польше Рудницким [Rudnicki et al., 1976] и в СССР И. Р. Рахимбаевым [1975, 1976].
Важность этих работ заключается в том, что они с полной очевидностью подтвердили концепцию гормональной регуляции ростовых процессов у луковичных культур.
Система гормональной регуляции роста луковичных растений представляется как совокупное взаимодействие эндогенных стимуляторных и ингибиторных факторов, главнейшими компонентами которых являются ауксины, ингибиторы, гиббереллин и цитокинины.
Накопленные сведения отражают роль отдельных гормонов в процессе морфогенеза и ростовых процессов.
Изучение состава эндогенных регуляторов роста, их накопления и распределения, изменения динамики их активности является важным для расширения представлений о гормональной регуляции покоя и роста у луковичных растений. Вместе с тем выяснение закономерностей эндогенной регуляции может служить основой для целенаправленного применения экзогенных искусственных физиологически активных веществ в управлении ростом и развитием растений.
Отрастание «покоящихся» луковиц — это замедленный, плавно протекающий процесс, который длится несколько месяцев. Выход луковиц из состояния «покоя» обеспечивается усиленным накоплением фитогормонов и одновременно с этим снятием тормозящего действия ингибиторов роста. Смещение баланса ингибиторы—фитогормоны происходит не внезапно, а последовательно путем «ступенчатой активации» фитогормонов. В самые начальные фазы покоя, когда внутри луковицы имеется недифференцированный конус нарастания, обнаруживаются только ауксины и цитокинины, затем содержание этих фитогормонов постепенно возрастает и достигает значительных величин, особенно в фазе дифференциации зачатков цветка. Существует очередность действия разных групп фитогормонов: вначале ауксины и цитокинины, затем гиббереллины.
Взаимосвязь между повышением гормональной активности и выходом луковиц из покоя наиболее отчетливо наблюдается при изучении эндогенных гиббереллинов. В самом начале периода покоя, когда конус нарастания еще не дифференцирован, не удается обнаружить гибберелловую активность. Лишь после формирования зачатков листьев удается выявить биологическую активность, характерную для гиббереллинов, как в апикальной почке, так и в запасающих чешуях.
Дальнейшее возрастание концентрации гиббереллинов (ГК) в апикальной почке происходит при формировании зачатка цветка. Перед выходом луковиц из покоя содержание ГК повышается в 20 раз по сравнению с начальными фазами покоя.
Значительную роль в переходе луковиц к отрастанию играют цитокинины [Рахимбаев, Соломина, 1975]. Они обнаружены еще в недифференцированном конусе нарастания, и их содержание повышается в процессе формирования листьев и цветка. Установлено, что цитокинины содержатся преимущественно в связанной форме в запасающих чешуях. При выходе из покоя наблюдается возрастание уровня свободных цитокининов.
Таким образом, в результате исследований фитогормонов и ингибиторов в связи с покоем и отрастанием луковичных растений выявлены следующие закономерности:
1) в процессе выхода луковиц из состояния покоя снижается содержание абсцизовой кислоты;
2) содержание фитогормонов (гиббереллина, индолин-уксусной кислоты, цитокипина) возрастает непосредственно перед выходом растений из покоя;
3) накопление фитогормонов обеспечивается преимущественно за счет их высвобождения из связанной формы.
Не менее убедительно физиолого-биохимическими исследованиями было показано, что в луковицах при пониженных положительных температурах происходит существенная перестройка системы эндогенных стимуляторов роста при заметном возрастании биологической активности цитокининов. Эндогенные гиббереллины при 22° находятся в связанном состоянии и лишь при охлаждении луковиц происходит резкое повышение активности [Aung, Hertogh, 1979].
В охлажденных луковицах накапливаются свободные формы гиббереллинов, обладающие высокой биологической активностью. В основе гормональных перестроек, происходящих под воздействием определенного периода охлаждения луковиц (конкретного для каждой культуры и даже сорта), лежит повышение уровня биологической активности цитокинина и гиббереллина, а также изменение места их локализации в растении.
Основные качественные и количественные изменения гиббереллина происходят в период активного синтеза и использования запасных веществ луковиц при укоренении и отрастании растений при низких положительных температурах и влажности, близкой к 100%.
Исследования позволили обнаружить в разных органах развивающихся луковиц тюльпанов около 20 эндогенных гиббереллинов. Наиболее активными (для тюльпанов) названы гибберелловые кислоты: А3, А4, А5, А7 [Aung et al., 1971].
Оунг и Рисе [Aung, Rees, 1974], исследуя биохимические изменения и образование предшественников гиббереллинов, пришли к выводу, что главный эффект эндогенных гиббереллинов — это стимуляция гидролитической энзиматической деятельности, активирующей превращение крахмала в запасающих чешуях луковиц.
Разные этапы роста луковичных растений, отличающиеся по скорости и характеру ростовых процессов (летний покой, выход из покоя, зимний покой, весеннее отрастание, интенсивный рост, торможение роста), регулируются соотношением и концентрацией фитогормонов и ингибиторов в разных частях растений в процессе вегетации. Каждая фаза роста характеризуется своеобразным гормональным статусом.
Как известно, многообразие физиологически активных веществ характеризуется одним общим свойством — они усиливают (как стимуляторы) или угнетают рост и жизнедеятельность растений [Рахимбаев, Соломина 1975].
Ростовые вещества косвенно влияют на течение процессов роста на определенных этапах, освобождая связанные фитогормоны и переводя их в активные формы.
Физиологически активные вещества участвуют в сложном процессе, который зависит от особенностей морфогенеза растения, времени стимуляции, концентрации, физиологически экзогенного используемого вещества и условий выращивания растений [Рахимбаев и др., 1976].
Выяснение закономерностей экзогенной регуляции у луковичных растений является основой для применения синтетических физиологически активных веществ.
У луковичных растений в естественных условиях (зимний период) и в условиях искусственного охлаждения и укоренения происходит накопление необходимого количества гиббереллинов, которые являются одной из главных причин выхода растений из состояния покоя и обеспечивают полноценный рост и цветение.
Первые работы по применению искусственных регуляторов роста в процессе выгонки луковичных растений появились сравнительно недавно. Като [Kato, 1966] отметил стимуляцию прорастания луковиц при обработке их гибберелловой кислотой А3.
В дальнейших исследованиях [Bragt, 1972; Aung et al., 1971] были испытаны различные формы разной концентрации гибберелловой кислоты (А3, А4+7) на охлажденных (в разной степени) и неохлажденных луковицах тюльпанов.
Полученные результаты показали, что введение экзогенного гиббереллина не может полностью заменить эффекты 12-недельного охлаждения (при температуре 5°). Кроме того, выявлено, что растения более чувствительны к действию экзогенного гиббереллина на ранних этапах развития, чем в период удлинения цветочного стебля.
В соответствии с установленными закономерностями были начаты опыты, связанные с использованием физиологически активных веществ в процессе выгонки, которые в ряде случаев дали положительные результаты [Былов и др. 1977; Смирнова, 1977]
В нашей работе исследовалась возможность сокращения периода охлаждения у тюльпанов, нарциссов, гиацинтов, мускари путем введения гибберелловой кислоты А3 в растения на разных этапах охлаждения. Проведено намачивание сухих охлажденных луковиц до посадки на выгонку 0,1%-ным раствором гиббереллина А3 (ГК—А3) в течение 24 и 48 ч.
Испытаны полив и опрыскивание растений 0,1%-ным раствором ГК—А3 в течение 12-недельного периода их укоренения и охлаждения (при 8°) после перемещения на свет.
Наибольший эффект отмечен в тех вариантах, когда предварительно охлажденные в течение 4 нед луковицы (при 8°) намачивали 48 ч в 0,1%-ном растворе ГК-А3 перед посадкой на выгонку. Отчетливо наблюдалось стимулирующее действие ГК-А3 на время зацветания - у тюльпанов раньше на 5-7 дней, у нарциссов - на 5, у гиацинтов — на 13 дней, чем в контроле, где луковицы намачивали в воде. У мускари отмечено цветение после обработки ГК-А3, в то время как в контроле оно отсутствовало (табл. 1).
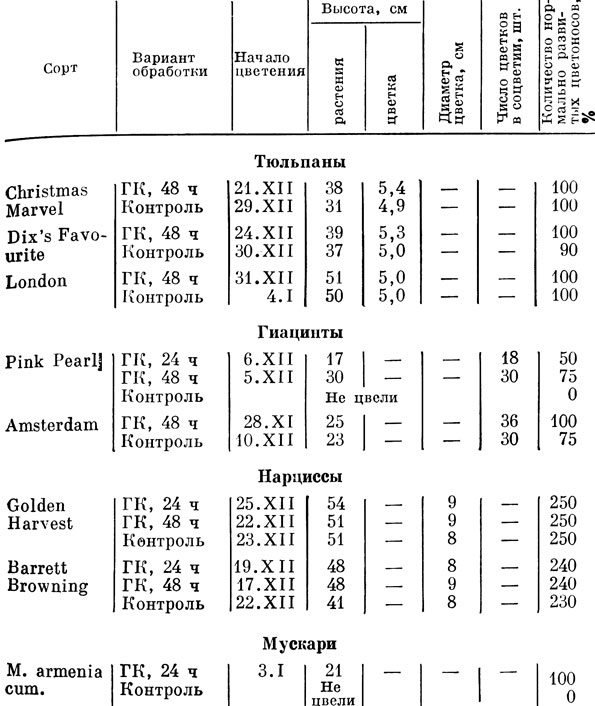
Таблица 1. Данные эффективности цветения тюльпанов, нарциссов, гиацинтов, мускари в ноябре, декабре при обработке ГК
При анализе действия экзогенных ростовых веществ на состояние покоя и рост луковичных растений обнаружена важная роль ингибиторов в этих процессах.
Шоубом и Хертогом [Shoub, Hertogh, 1974] изучена возможность изменения высоты цветочного стебля тюльпанов при внесении в почву ГК—А3 совместно с анцимидолом при посадке частично охлажденных луковиц на выгонку.
Исследования показали эффективность удлинения цветочного стебля при внесении 25 мг ГК—А3 на 1 растение и сокращение высоты цветоноса до 10 см возможно при внесении 25 мг анцимидола на 1 растение.
Среди экзогенных физиологических активных веществ ретандаят хлорхолинхлорид обладает ингибиторным действием и тормозит рост стебля в длину, стимулирует его утолщение, влияет на цветение растений. Тюльпаны, обработанные хлорхолинхлоридом после 12 нед охлаждения при 5°, цветут позже, чем в контроле [Read el al., 1974].
Выявляется важная закономерность, что только при определенном соотношении фитогормонов и ингибиторов возможен переход от одного физиологичского состояния луковиц к другому. Ни один из гормонов не может самостоятельно вызвать покой или выход из покоя.
В связи с этим мы полагаем, что по мере углубления наших знаний о влиянии отдельных факторов на физиологические процессы, протекающие на определенных этапах морфогенеза луковичных растений, значительно расширятся наши возможности для разработки в полном смысле управляемой культуры, обеспечивающей получение цветущих растений в заранее планируемые сроки.
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://flowerlib.ru/ 'Библиотека по цветоводству'