ГЛАВА VI: ВЛИЯНИЕ РОДИОЛЫ НА ЦЕНТРАЛЬНУЮ НЕРВНУЮ СИСТЕМУ (Глава IV написана совместно с Т. Ф. Мариной.)
Для характеристики психостимуляторов значительный интерес представляет изучение влияния их на ЦНС. Соответствующие экспериментальные и клинико-физиологические исследования, выполненные в нашей лаборатории (А. С. Саратиков и соавт., 1965; Т. Ф. Марина, 1966,1968; И. М. Калико и А. И. Тарасова, 1965, 1966), выявили сходство в действии препаратов родиолы, элеутерококка, женьшеня и левзеи на функциональное состояние головного и спинного мозга.
В хронических опытах на 47 кроликах с вживленными электродами Т. Ф. Марина и Л. П. Алексеева (1968) исследовали влияние экстракта родиолы, родозина и салидрозида на фоновую биоэлектрическую активность сенсомоторной и затылочной областей коры головного мозга, а также на реакцию активации, получаемую в ответ на раздражение седалищного нерва или звуковое раздражение, и усвоение ритма световых мельканий.
Все изученные препараты родиолы оказывали однотипное действие на электроэнцефалограмму (ЭЭГ), которое в значительной мере зависело от состояния фоновой активности коры. В большинстве опытов фоновая электрокортикограмма интактных животных характеризовалась «ритмом покоя» и лишь у некоторых кроликов наблюдалась либо непрерывная десинхронизация, либо чередование участков десинхронизации с участками «ритма покоя».
Действие родиолы более отчетливо проявлялось при первом типе биоэлектрической активности. Через 15—30 минут после подкожного и через 5—10 минут после внутривенного введения препаратов наступала активация ЭЭГ: исчезали высокоамплитудные медленные колебания, сонные веретена; В зрительных областях коры появлялся синхронизированный ритм частотой 5—6 кол/сек., в сенсомоторных — низкоамплитудная быстрая активность. Аналогичное усиление спонтапной биоэлектрической активности выявлено и в подкорковых структурах — хвостатых ядрах, миндалине, дорсальном гиппокампе, переднем и заднем гипоталамусе, неспецифических ядрах таламуса, ретикулярной формации среднего мозга (рис. 17) (Т. Ф.Марина и Л. П.Алексеева, 1971).
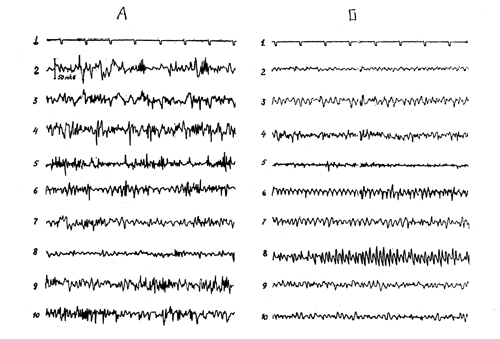
Рис. 17. Влияние салидрозида (4 мг/кг внутревенно) на ЭЭГ кролика. А - до введения, Б - через 15 минут после введения препарата. Сверху вниз - отметка времени; 2, 3 - сенсомоторная и зрительная области коры, 4 - хвостовое ядро, 5 - миндалина, 6 - ретикулярная формация среднего мозга, 7 - неспецифические ядра таламуса; 8, 9, 10 - передний, задний и латеральный гипоталамус.
Как правило, препараты родиолы вызывали не непрерывную активацию (что характерно для действия фенамина и I пиридрола), а чередование участков ЭЭГ активации с отдельными группами более медленных волн и редуцированных веретен в течение 45—60 минут.
У животных с исходным активным фоном действие родозина и салидрозида проявлялось слабее и характеризовалось укорочением или исчезновением участков ритма «покоя» и учащением низкоамплитудной синхронизированной ритмики в зрительных областях коры.
Активирующее действие препаратов родиолы на фоновую биоэлектрическую активность сопровождалось изменениями функциональных проб, свидетельствующими о повышении возбудимости и лабильности нервных клеток головного мозга: усиливалась и удлинялась реакция ЭЭГ активации в ответ на электро- и фоностимуляцию без сдвига порога раздражения; улучшалась реакция следования ритму световых мельканий, что проявлялось в увеличении индекса усвоения, небольшом расширении диапазона усваиваемых частот, в иррадиации реакции следования на сенсомоторные области коры (рис. 18).
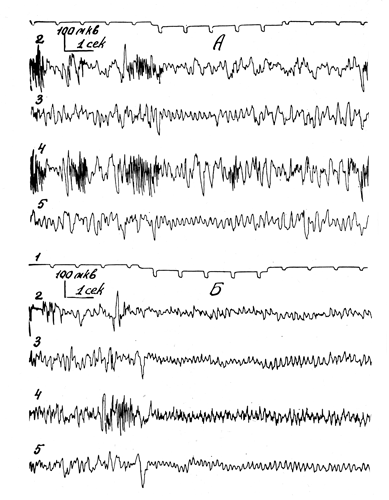
Рис. 18. Влияние салидрозида (4 мг/кг внутривенно) на реакцию активации, вызванную электрическим раздрожением седалищного нерва. А - до введения, Б - через 10 минут после введения препарата. Сверху вниз: 1 - отметка времени; 2, 4 - правая и левая сенсомоторные; 3, 5 - правая и левая затылочные области коры.
Таким образом, препараты родиолы вызывают превалирование возбудительного процесса в коре головного мозга, повышают возбудимость и лабильность нервных клеток.
Следует отметить, что наиболее четко указанные изменения ЭЭГ были выражены при введении кроликам 0,2 мл/кг родозина и 2 мг/кг салидрозида. Увеличение дозы родознна до 1—2 мл/кг и родиолозида до 10—20 мг/кг не только не сопровождалось усилением десинхронизирующего эффекта, а влекло за собой появление медленной активности с уменьшением реакции ЭЭГ активации в ответ на электро- и фоностимуляцию и ухудшением реакции следования (рис. 19). Подобный сдвиг ЭЭГ показателей соответствует тормозному состоянию исследуемых структур мозга.
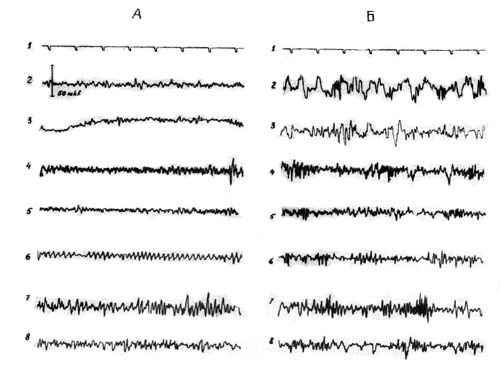
Рис. 19. Влияние салидрозида (20 мг/кг внутривенно) на ЭЭГ кролика. Обозначения те же, что на рис. 17.
Препараты родиолы не предотвращают и не прерывают депримирующего действия на электрокортикограмму хлоралгидрата (75 мг/кг внутривенно) и барбитал-натрия (100 мг/кг внутривенно), а лишь несколько уменьшают его интенсивность и способствуют более скорой нормализации ЭЭГ. Они уменьшают степень и длительность синхронизирующего действия центрального м-холинолитика бензацина (0,1 мг/кг в/в).
Введение салпдрозида в боковые желудочки головного мозга вызывало изменения ЭЭГ, аналогичные вышеописанным, при значительно меньших дозах препарата: 0,2—0,4 мг/кг проявляли десинхронизирующий эффект; 0,8—1,0 мг/кг в части опытов вызывали появление медленной высокоамплитудной активности. Это может свидетельствовать о наличии у салидрозида прямого центрального действия.
Для выяснения вопроса, с каким отделом головного мозга связаны ЭЭГ эффекты родиолы, Т. Ф. Марина (1968) исследовала влияние родозина и салидрозида на биоэлектрическую активность коры головного мозга при различной степени изоляции ее от стволовых структур.
Перерезки мозга кроликов производились на следующих уровнях: между первым и вторым шейными позвонками (препарат «encephale isole»), трягеминальном (каудальнее задних бугров четверохолмия и тотчас перед выходом тройничных нервов), межколликулярноv (между буграми четверохолмия, впереди варолиева моста), премезенцефаличееком (впереди передних бугров четверохолмия позади мамиллярных тел).
Активирующий эффект препаратов родиолы на ЭКоГ сохранялся на препарате ensphale isole, исключающем всю периферическую импульсацию, кроме трех пар черепномозговых нервов, но выражен был слабее, чем у животных с интактным мозгом: активация фоновой биоэлектрической активности наблюдалась в 57%, усиление ответной реакции на звуковое раздражение в 50% и улучшение реакции следования в 83% опытов. У интактных животных перечисленные изменения ЭКоГ зарегистрированы соответственно в 78%, 87% и 100% опытов.
На препарате мозга с тригеминальным сечением, для которого характерна реакция активации, на электрокортикограмме вследствие разобщения коры с синхронизирующими механизмами каудального отдела ретикулярной формации отчетливо проявлялся синхронизирующий эффект больших доз родозина (1 мл/кг) и салидрозида (10 мг/кг).
При межколликулярном сечении (cerveau isole), разобщающем кору с каудальными отделами ретикулярной формации и характеризующемся наличием на электрокортикограмме медленных волн различной амплитуды, чередующихся с веретенами, препараты родиолы либо не оказывали активирующего влияния на биоэлектрическую активность коры, либо способствовали появлению коротких участков тета-ритма (частотой 4—5 кол/сек.) в теменных и затылочных областях коры, незначительному улучшению реакции следования, наслоению частого ритма на высокоамплитудные медленные волны. Синхронизирующий эффект больших доз проявлялся при этом уровне сечения достаточно отчетливо в виде еще большего замедления ритма колебаний.
Наконец, на препарате мозга с премезенцефалическим сечением, отделяющим весь средний мозг от вышележащих отделов головного мозга, родозин и салидрозид не оказывали влияния на ЭЭГ.
Таким образом, опыты с сечением мозга свидетельствуют, что для проявления активирующего влияния препаратов родиолы на ЭКоГ необходимы два условия: во-первых, сохранение связи каудальных отделов ретикулярной формации с корой головного мозга; во-вторых, наличие афферентной импульсации. Синхронизирующий эффект на ЭКоГ больших доз родозина проявляется как при каудальных, так и при более ростральных сечениях мозгового ствола.
Для выяснения характера действия препаратов золотого корня на функциональное состояние различных отделов головного мозга Т. Ф. Мариной и Т. М. Плотниковой (1973) исследовано влияние родозина на электрографические реакции коры больших полушарий и некоторых подкорковых структур, получаемые путем их электростимуляции. В 119 хронических опытах на 22 ненаркотизированных кроликах электростимуляцию сенсомоторной коры, дорсального гиппокампа, базола-теральных ядер миндалевидного комплекса, неспецифических ядер таламуса и мезенцефалической ретикулярной формации производили через биполярные электроды в стеклянной изоляции с помощью электронного стимулятора ЭСТ-10. Показателями функционального состояния изучаемых структур являлись следующие электрографические реакции: для сенсомоторной коры, дорсального гиппокампа и ядер миндалины — порог и длительность судорожных разрядов последействия; для мезенцефалической ретикулярной формации — порог и длительность реакции активации; для неспефицической тала-мокортикальной системы — порог и амплитуда корковых ответов реакции вовлечения.
Анализ проведенных опытов показал, что под влиянием малой дозы родозина, проявляющей мягкое активирующее влияние на спонтанную биоэлектрическую активность головного мозга, наблюдается преимущественно изменение функционального состояния структур ретикуло-таламокортикальной системы. Так, родозин в дозе 0,2 мл/кг в течение 1 часа статистически достоверно удлинял реакцию активации, возникавшую в ответ на электростимуляцию мезенцефалической ретикулярной формации, существенно не влияя па величину ее порога; проявлял тенденцию к повышению порога реакции вовлечения через 30 минут — 1 час после введения. При этом не нарушался процесс торможения в коре больших полушарий, так как судорожный порог этой структуры повышался на 0,25±0,08 в.
Учитывая, что активирующий эффект препаратов родиолы на ЭКоГ проявляется лишь при сохранении связей ретикулярной формации с корой головного мозга, а сдвиг в уровне вэзбудимости неспецифической таламокортикальной системы появляется позднее, чем в мезенцефалической ретикулярной формации, можно полагать, что первичным эффектом родозина является воздействие препарата на восходящую ретикулярную активирующую систему. Снижение возбудимости таламокортикальной рекруитирующей системы развивается, по-видимому, вторично.
Поскольку препараты родиолы почти не изменяют порога реакции активации в ответ на электростимуляцию ретикулярной формации среднего мозга и активирующее влияние их на кору больших полушарий уменьшается с выключением большей части афферентной импульсации, создается впечатление, что они не возбуждают ретикулярную формацию непосредственно, а повышают ее чувствительность к внешним влияниям, поступающим в эту структуру через коллатерали. Этим они существенно отличаются от стимуляторов группы фенамина, которые обладают прямым возбуждающим действием на донто-мезенцефалическую ретикулярную формацию.
В дозе 1 мл/кг, проявляющей адаптогенное действие (см. гл. VI) и синхронизирующий эффект на ЭЭГ, родозин укорачивал реакцию активации и в течение 1 часа статистически значимо повышал ее порг на 0,1 ±0,04 в, увеличивал амплитуду корковых ответов реакции вовлечения в среднем в 1,5 раза, вызывал удлинение этой реакции и достоверное понижение ее порога на 0,14±0,06 в на протяжении двух часов наблюдения (рис. 20). Таким образом, успокаивающее влияние на биоэлектрическую активность мозга адаптогенной дозы родозина обусловлено не только угнетением мезенцефалической ретикулярной формации, но и активацией неспецифической таламокортикальной системы. Интересно, что при этой дозе препарата отчетливо проявлялось его влияние на функциональное состояние лимбических структур: наблюдалось снижение судорожного порога в дорсальном гиппокампе и повышение его в ядрах миндалевидного комплекса.
Т. Ф. Мариной и соавторами (1971, 1973) исследовано влияние препаратов родиолы розовой на адренергические процессы в ЦНС путем изучения взаимодействия родозина и салидрозида с фенамином — веществом, вызывающим преимущественно центральное адренергическое возбуждение (Р. Ю. Ильюченок, 1965; П. А. Шаров, 1967; Glowinsky, Snyder, Axelrod, 1966 и др.). и аминазином, проявляющим свойства центрального адренолитика (Ф. Б. Брэдли, 1962).
Опыты поставлены на 110 мышах, 34 крысах и 9 кроликах. Изучено влияние родозина и салидрозида на фенаминовое двигательное возбуждение у мышей (путем регистрации вертикального компонента ориентировочной реакции животных — «вставаний»), токсичность фенамина у изолированных и сгруппированных мышей, фенаминовую гипертермию и стереотипию у крыс. Препараты родиолы вводили мышам и крысам подкожно за 30 минут до фенамина. Взаимодействие препаратов родиолы с аминазином (1 и 3 мг/кг внутривенно) исследовано методом ЭЭГ в хронических опытах на кроликах с электродами, вживленными в сенсомоторную, зрительную области коры больших полушарий и основные подкорковые структуры. Салндрозид и родозин вводили внутривенно за 15 минут до инъекции аминазина или на 15-ой минуте его действия.
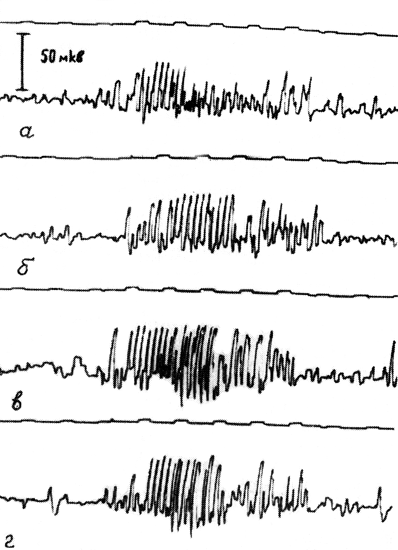
Рис. 20. Влияние родозина (1 мл/кг внутривенно) на амплитуду корковых ответов реакции вовлечения, вызванной раздражением неспецифических ядер таламуса. Верхняя кривая - отметка времени и раздражения, нижняя кривая - ЭЭГ сенсомоторной области коры; а - контроль; б, в, г, - спустя 30, 60 и 90 минут после введения родозина.
Салидрозид в дозе 10 мг/кг статистически достоверно удлинял, а в дозе 30 мг/кг ускорял наступление, усиливал и удлинял фенаминовую гиперактивность животных; при дозе 100 мг/кг препарат не оказывал влияния на этот эффект фенамина. Препарат (30 и 100 мг/кг) не влиял на токсичность фенамина у изолированных мышей и несколько усиливал гибель животных от фенамина при групповом их содержании. Родозин при индивидуальном применении не изменял ректальную температуру крыс в течение 4—6 часов наблюдения. Родозин (2 и 10 мл/кг) и салидрозид (30 мг/кг) предупреждали гипертермический эффект фенамина. Введение родозина (0,4 мг/кг) не влияло на длительность фенаминовой стереотипии у крыс, при дозах 2 и 10 мл/кг препарат статистически значимо укорачивал продолжительность этого феномена.
Препараты родиолы розовой как при профилактическом, так и лечебном введении, не изменяли длительности депримирующего действия аминазина на биоэлектрическую активность головного мозга кроликов и лишь незначительно уменьшали степень угнетающего эффекта аминазина по показателям фото- и фоностимуляции.
Таким образом, препараты золотого корня в малых и средних дозах проявляют адренопозитивное действие, усиливая фекалиновую гиперактивность и токсичность фенамина у сгруппированных мышей, уменьшая интенсивность угнетающего воздействия аминазина на ЭЭГ кроликов. В средних и больших дозах в опытах на крысах они обнаруживают адреноне-гатнвные свойства, предупреждая развитие фенаминовой гипертермии и укорачивая длительность фенаминовой стереотипии. Обращает на себя внимание большая в ыраженностьтормозящего действия препаратов родиолы на центральные эффекты фенамина у крыс, а усиливающего — у мышей. Эти различия могут быть обусловлены различной видовой чувствительностью животных к препаратам родиолы и особенностями биотрансформации фенамина в организме использованных видов животных.
Описанный выше антагонизм препаратов родиолы и м-холинолитика бензацина побудил Т. Ф. Марину (1973) исследовать влияние салидрозида на холинергические процессы в ЦНС. В опытах на 190 белых мышах ею изучено взаимодействие салидрозида с веществами, возбуждающими и блокирующими холинорецепторы ЦНС. В качестве центральных м- и н-холиноблокаторов использовали соответственно амизил (5 мг/кг подкожно) и спазмолитин (30 мг/кг подкожно), а м- и н-холиномиметиков — ареколин (25 мг/кг подкожно) и никотин (12 мг/кг подкожно). В опытах с х'олиноблокаторами учитывали в качестве показателя двигательной активности количество подъемов животного на задние лапы до введения исследуемых препаратов и через 30, 60 и 120 минут после введения. В экспериментах с ареколином и никотином интенсивность судорог регистрировали при помощи актометра. Салидрозид инъецировали подкожно в дозах 10, 30 и 100 мг/кг за 30 минут до введения холинергических веществ.
Как видно из табл. 22, салидрозид в дозах 10 и 30 мг/кг существенно увеличивал интенсивность никотинового тремора у мышей. Гибель животных при этом не только не увеличивалась, но даже несколько снижалась (с 25—28,5 до 13,8—16,7%). Таблица 22 Влияние салидрозида на интенсивность никотиновых судорог у мышей.
| Группы животных | Показ актометра | ||||||||||
| Контроль | Салидрозид в мг|кг | ||||||||||
| 10 | 30 | 100 | |||||||||
| n | M±m | n | M±m | P | n | M±m | P | n | M±m | P | |
| I II III |
15 10 7 |
78,7±14,8 43,0±15,8 180,0±40,9 |
13 9 8 |
141,7±25,8 82,5±25,8 644,7±207,5 |
0,04 0,02 0,04 |
13 9 10 |
170,3±42,2 118,3±24,8 560,3±160,6 |
0,05 0,02 0,04 |
20 9 - |
112,8±16,2 126,3±79,0 - |
0,12 0,08 - |
(Примечание: n - количество животных в группе; группы отличаются друг от друга весом мышей и чувствительностью актометра. )
Салидрозид в дозе 10 мг/кг не влиял на интенсивность ареколинового тремора, а в дозах 30 и 100 мг/кг — уменьшал его. Так, в контрольных опытах этой серии показания актометра составляли 291 ±72; в экспериментах с 30 до 100 мг/кг салидрозида соответственно 84+34 (Р = 0,02) и 96±29 (Р = 0,03). Препарат уменьшал степень угнетающего влияния на двигательную активность мышей центральных холиноблокаторов амизила и спазмолитика.
Таким образом, препараты родиолы обладают центральным н-холинопозитивным и в больших дозах центральным м-холи-нонегативным действием.
Влияние препаратов родиолы на функциональное состояние спинного мозга (Т. Ф. Марина, 1966) изучено путем измв рения электровозбудимости лягушек по методу Scheminsky (1924) в модификации С. Я. Арбузова (1960) и скрытого времени ипсилатерального сгибательного рефлекса задней конечности кролика по В. В. Закусову (1948).
Опыты по определению электровозбудимости лягушек за ключались в нахождении порога переменного тока, вызывающего у животных, помещенных в сосуд с водой, тетанические судороги. Измерения производились каждые 20 минут в течение 3 часов после введения исследуемых препаратов. Эксперименты выполнены на интактных животных, на фоне действий веществ, угнетающих центральную нервную систему (хлоралгидрат, барбитал-натрий), и в условиях перерезки спинногШ мозга у кроликов на уровне 9—10 грудных позвонков.
Экстракт родиолы (0,05; 0,2; 1 мл/кг подкожно) не влиял на электровозбудимость интактных лягушек. Однако, введенный в эффективных дозах (0,2; 1 мл/кг) животному на высота депримирующего действия хлоралгидрата (400 мг/кг) препарат способствовал более ранней нормализации исследуемой показателя. После децеребрации животных этот эффект родиолы утрачивался.
Влияние экстракта родиолы на скрытый период флексор ного рефлекса (СПФР) задней конечности изучено на 21 кролике. Измерение СПФР производили с помощью рефлексометра типа РФ-1-55 через каждые 5 минут в течение 1,5—2 часов Экстракт родиолы в тех ж«е дозах у большинства интакт ных кроликов не вызывал статистически достоверных откло нений от нормы СПФР. Только 1 кролик из 8 реагировал нг введение экстракта (1 мл/кг) укорочением СПФР на 25%. Этот эффект проявлялся через 45 минут после введения препарата и сохранялся в течение 75 минут. Экстракт родиолы: введенный за 1 час до внутривенной инъекции хлоралгидрата (75 мг/кг), статистически значимо ускорял нормализацию СПФР. После перерезки спинного мозга у кроликов на уровне 9—10 грудных позвонков нормализующее влияние родиолы на исследуемый показатель не проявлялось.
Следовательно, способность родиолы улучшать межнейроннуго передачу возбуждения в спинном мозгу обусловлена воздействием его на высшие отделы ЦНС и, возможно, осуществляется посредством усиления нисходящих активирующих влияний ретикулярной формации.
Наблюдения над 30 здоровыми лицами и 45 больными неврозами (И. М. Калико, А. А. Тарасова, 1965, 1966), включающие исследования высшей нервной деятельности по речедвигательной методике А. Г. Иванова-Смоленского и по методике ассоциативного эксперимента, показали, что курсовое назначение! экстракта родиолы (по 10 капель 3 раза в день в течение 10 дней) у здоровых людей повышало внимание, память и силу возбудительного процесса. У больных неврозами после однократного и в особенности курсового приема препарата наступало усиление возбудительного и тормозного процессов и нормализация их подвижности (см. гл. VII).
| Условия опыта | Время в сек. | Количество | |||
| пробега по лабиринту | пребывания на площадке | подъема по канату | пересеченных линий | подъемов на задние лапки | Контроль Элеутерозид Рк Контроль Родиола Рк |
16,8±0,5 14,2±1,2 0,067 12,0±0,8 8,7±1,0 0,021 |
14,7±2,0 15,0±0,5 0,92 8,5±1,0 6,0±1,2 0,13 |
6,8±0,6 5,5±0,7 0,18 6,0±0,6 3,6±0,5 0,009 |
46±4 38±4 0,18 23±3 35±4 0,032 |
18±2 7,8±1 0,001 18±7 18±1 1,0 |
С этими наблюдениями согласуются результаты исследования влияния настоя корневищ родиолы розовой (0,5 мл/100 г веса) на некоторые показатели функционального состояния ЦНС крыс, полученные в лаборатории проф. К. А. Мещерской (Г. Ампилогова и Н. Присяжнюк). Опыты проводили на заранее тренированных беспородных крысах, используя станок Винтера и Флатекера и лабиринт. Как видно из табл. 23, в отличие от суммы элеутерозидов (5 мг/100 г) препарат родиолы существенно сократил время пробега по лабиринту и почти вдвое ускорил подъем животных по канату, что свидетельствует о стимулирующем условнорефлекторную деятельность влиянии препаратов родиолы; возрос также и горизонтальный компонент ориентировочного рефлекса.
Влияние суммы элеутерозидов и настоя родиолы на некоторые показатели состояния ЦНС крыс (средние из 7-8 опытов)
|
ПОИСК:
|
При использовании материалов активная ссылка обязательна:
http://flowerlib.ru/ 'Библиотека по цветоводству'